

今回は、チコちゃんに叱られる! ▽関東と関西▽ナマケモノの不思議▽焼き肉の謎 初回放送日:2024年9月20日を紹介。
なんでナマケモノはなまけているの?


なんでナマケモノはなまけているの?
チコちゃん「ねぇねぇ岡村、この中で一番、休みの日はのんびり過ごす余裕のある大人ってだーれ?」
チコちゃん「なんでナマケモノはなまけているの?」
チコちゃんの答えは、「なまけないと生きていけないから。」
なまけないと生きていけないから
ナマケモノは中央アメリカから南アメリカの熱帯雨林に生息。
木の上で葉っぱなどを食べながら生活する動物。
ほとんど動かない様子が名前の由来。
ナマケモノにはフタユビナマケモノとミユビナマケモノの2種類。


特に動かないとされているのはミユビナマケモノ。
ナマケモノの祖先はもともと地上で生活していたといわれています。
ある時期に木の上で葉っぱを食べて暮らす種類が登場。
なぜ木に登ったのか?いつ頃そうなったのか?のは、はっきりしていません。
樹上生活に適応した種類が現在のナマケモノに繋がっていると考えられています。
草食動物にとって木の上で葉っぱを食べて生きるのはかなり大変。
サバイバル能力としてナマケモノが生み出したのがなまける事でした。
葉っぱや草はカロリーが低く、かつ消化しづらいので大量に食べる必要があります。
生きるために大食いしないといけないとなると体は必然的に大きく重くなります。
そうなると枝にぶら下がったり、木に登ることに不都合になります。
そこでナマケモノは沢山食べなくても生きていけるように超省エネボディーに進化。
「とにかく食べない。食べないから動かない。」が基本。
ナマケモノの一日の食事量は葉っぱ約200g程度。
キリンの場合は一日58kgほど食べます。


体重あたりの食事量で比較すると、ナマケモノの体重、約4.3kg対して200gだと4.5%。
キリンの体重は、761kgで58kgなので7.6%。
通常は体重が軽くなるほど体重あたりの食べる量は増えるのが普通。
ナマケモノは体重で見ても超少食。
それだけ食べないので全く動かないナマケモノ。
一日のうち8割以上にあたる19時間は何もせずにひたすらじっとしています。

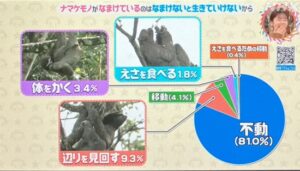
19時間、ほぼ動かないナマケモノ。
また、体に藻が生えているというナマケモノはもはや木と同化している状態。


なまける生活は敵から身を守る重要な役割。
ギリギリのエネルギー戦略で日々を暮らしているナマケモノ。
天気の悪い日が続いてしまうと体温が下がり過ぎて命の危機に瀕してしまうという弱点も。
また、大体1週間に1回はトイレタイムが。
その時には木から下ります。
そのタイミングは敵に襲われる可能性が高くなります。
ゆっくり木を下りて、上半身は木にしがみついたままおトイレ。
この時に外敵に襲われてもナマケモノは素早く動くことができません。
木の上でトイレをしない理由は、はっきり分かっていないとのこと。
コアラも木の上でじっと動かずに一日の大半を過ごす動物。
それは主食のユーカリの解毒や消化に体力が必要なために省エネ生活をしていると異なる理由。


最後に進化しすぎと困ったことになっている動物を紹介。
一つ目が、日本にやってくる渡り鳥のオオハム。


水中を泳いだりもぐったりできますが、地上で歩くのはうまく歩けなくなりました。
2つ目が、ロシアや北欧で生息するヘラジカ。
角が大きいオスが縄張り争いに勝つそうですが、大きくなりすぎて絡まってケンカができない状態に。

結論
というわけで、
「なんでナマケモノはなまけているの?」は、
「なまけないと生きていけないから」
でした。
解説してくれたのは
奈良教育大学の村松大輔特任准教授。
職名・氏名
[特任准教授] 村松 大輔 (MURAMATSU Daisuke)
連絡先 TEL:0742-27-9142
MAIL:muramatsu.daisuke.du
最終学歴 京都大学理学研究科博士後期課程修了 (2009年) 博士(理学)(京都大学 2009年)
所属 自然環境教育センター
専門分野 動物行動学・生態学・行動生態学
主な担当授業
自然環境学
生態学実験
生活
小学校生活
臨海実習
主な業績
Muramatsu D, Vidal LV, Costa ER, Yoda K, Yabe T, Gordo M (2022) Low-cost thermoregulation of wild sloths revealed by heart rate and temperature loggers. Journal of Thermal Biology, 110:103387. https://doi.org/10.1016/j.jtherbio.2022.103387
Hirakawa H, Muramatsu D, Gordo M, Takii A, Izumiyama S (2022) GAP method: a gap-based technique for outlier detection and its application to wildlife GPS data. Research Square, https://doi.org/10.21203/rs.3.rs-1567713/v1
Tina FW, Muramatsu D (2022) Males signal their breeding burrow characteristics to females in the fiddler crab Austruca perplexa. Behaviour, 159:1045–1062. https://doi.org/10.1163/1568539X-bja10165
Muramatsu D (2021) Sand-bubbler crabs distinguish fiddler crab signals to predict intruders. Behavioral Ecology and Sociobiology, 75:125.https://doi.org/10.1007/s00265-021-03066-5
Tina FW, Muramatsu D (2021) Small males of the fiddler crab Austruca perplexa court more in the off-peak breeding period when large males court less. Journal of Ethology, 39:297–308. https://doi.org/10.1007/s10164-021-00703-4
王子 修, 石田正樹, 村松大輔, 和田恵次 (2020) 体サイズ・密度・季節変化からみたチゴガニ雄のwaving頻度.Cancer, 29: 19–24. https://doi.org/10.18988/cancer.29.0_19
村松 大輔 (2020) 自然教育とナマケモノ. 自然と教育, 30: 15–19. http://hdl.handle.net/10105/00013356
Tina FW, Muramatsu D (2020) Size-dependent mating preference of the male fiddler crab Austruca perplexa. Ethology, 126(1): 68–75. https://doi.org/10.1111/eth.12961
Muramatsu D (2017) Sand ornaments used to attract females are avoided by rival males in the fiddler crab Uca lactea. Ethology Ecology & Evolution, 29(5): 501–510. https://doi.org/10.1080/03949370.2016.1243156
Muramatsu D, Koga T (2016) Fighting with an unreliable weapon: opponent choice and risk avoidance in fiddler crab contests. Behavioral Ecology and Sociobiology, 70(5): 713–724. https://doi.org/10.1007/s00265-016-2094-2
Itoh N, Muramatsu D (2015) Patterns and trends in fruiting phenology: Some implications for important chimpanzee diet. In: Mahale Chimpanzees: 50 Years of Research. Cambridge, Cambridge University Press. pp.174–194
Muramatsu D (2011b) The function of the four types of waving display in Uca lactea: Effects of audience, sand structure, and body size. Ethology 117(5): 408–415. https://doi.org/10.1111/j.1439-0310.2011.01884.x
Muramatsu D (2011a) For whom the male waves: Four types of claw-waving display and their audiences in the fiddler crab, Uca lactea. Journal of Ethology 29(1): 3–8. https://doi.org/10.1007/s10164-010-0216-2
Muramatsu D (2010b) Temporary loss of a sexual signal (claw loss) affects the frequency of sand structure construction in the fiddler crab, Uca lactea. Ethology Ecology & Evolution 22(1): 87–93. https://doi.org/10.1080/03949370903516040
Muramatsu D (2010a) Sand structure construction in Uca lactea (De Haan, 1835) is related to tidal cycle but not to male or female densities. Crustaceana 83(1): 29–37. https://doi.org/10.1163/001121609X12487811051624
Muramatsu D (2009) To build or not to build—or to destroy burrow hoods in a population of Uca lactea. Journal of Crustacean Biology 29(3): 290–292. https://doi.org/10.1651/08-3122.1
研究と教育について
カニの一種にみられる様々なハサミ振りディスプレイの意味や、闘争の際に行われるオスどうしの駆け引き、巣穴の周りに作られる奇妙な構造物の機能など、視覚シグナルに関係する動物の行動や戦略について研究しています。
また、ブラジルのナマケモノを対象に、省エネルギー戦略や活動パターン、採餌生態などの研究を行っています。
所属学会
日本動物行動学会
Animal Behavior Society
講演のテーマ
「あなたの知らないナマの世界」 2022年
「ナマケモノのふしぎ:心拍と体温から探る怠け方の極意」 2021年
「スリナム共和国ナマケモノ保護センターへオンライン訪問」 2021年
「がんばらない生き方:ナマケモノの不思議なスローライフ」 2020年
「ナマケモノは なぜ、 どのように怠けているのか」 2020年
「心拍と体温から探るナマケモノの省エネ生活」 2019年
個人のページ https://www.researchgate.net/profile/Daisuke-Muramatsu
個人のページ2 https://researchmap.jp/preguica
(大学HPより)
今回も最後まで読んでくれてありがとう。
他の記事もよろしくね。
過去放送日別一覧ページ